
Human Pre-History and the Making of the Races, Part 2: Genetic Distances
Confirmation by modern genetic studies of the traditional racial classification categories
What are the percentages of genetic differences between the human races, indicating their relationships? Perhaps the best global scale study to date on this subject is still that of Masatoshi Nei and Arun K. Roychoudhury from Evolutionary Relationships of Human Populations on a Global Scale (1993). Subsequent studies, which have included increasing numbers of alleles but have usually been regional rather than global in scale, have been consistent with Nei and Roychoudhury’s results. The following table (Fig. 1 below) of estimates of genetic differences between human populations is from their study.
The following table of percentages of genetic differences between human populations presents the estimates for 19 populations from the above table in an easier to read and understand format. The human-chimpanzee genetic difference, giving the greatest degree of difference from the commonly accepted range using the same methodology, is added for context and comparison.
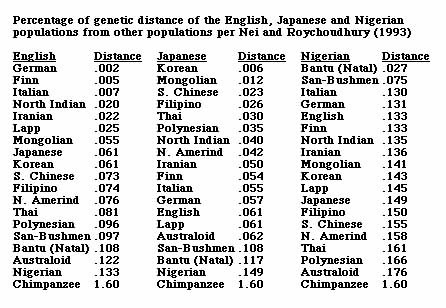
If one were to spatially visualize the first column of the above scale, with a German standing at a distance of 20 feet from an Englishman, a Finn would stand at a distance of 50 feet, an Italian at 70 feet, a northern Indian at 200 feet, a Japanese at 610 feet, a North American Amerindian at 760 feet, a Nigerian at 1,330 feet, and a Chimpanzee at 16,000 feet. The greatest percentage of genetic difference is .176% between Nigerians and Australian Aborigines. This is 11% of the genetic difference of 1.6% between humans and chimpanzees, different biological Families whose ancestral lines are believed to have separated about 5.5 million years ago. The .133% genetic difference between the English and Nigerian populations is 8.3% as large as the genetic difference between humans and chimpanzees. The .061% genetic difference between the English and Japanese or Korean populations is 3.8% as large as the genetic difference between humans and chimpanzees. Seen in this context, these are very significant genetic differences. It is also worth noting that for both the English and the Japanese, representing Europeans and Northeast Asians, the greatest percentage of genetic difference is with the Nigerians, and that the degree of this difference, .133% for the English and .149% for the Japanese, is very similar. By comparison, the English and Japanese degree of difference from the Australian Aborigine population, .122% for the English and .062% for the Japanese, is very different, with the English-Australoid difference twice as great as the Japanese-Australoid difference, presumably explained by the more recent divergence of the Mongoloid and Australoid branches and the extent of their common Denisovan mixture (see above).
The phylogenetic tree below, presenting a visual schematic of the genetic relationship between the different human populations, is also from the 1993 study by Nei and Roychoudhury.
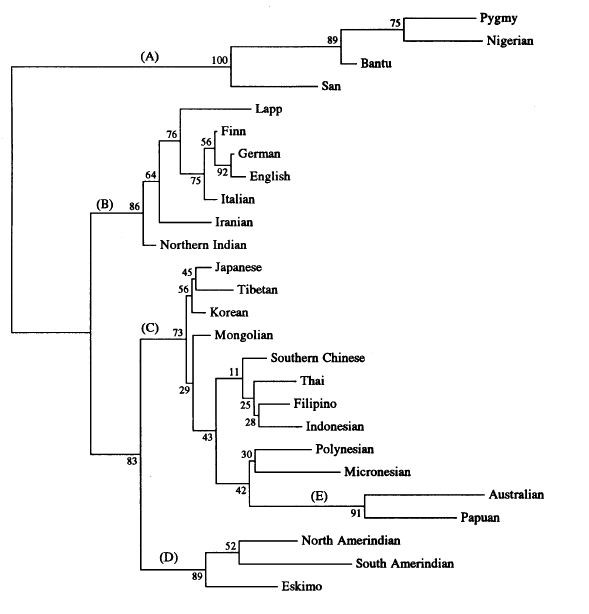 As seen in the above phylogenetic tree the major genetic divisions or branches of human populations are Africans (A), Caucasians (B), Greater Asians (C), Amerindians (D) and Australopapuans (E). Genetic studies have consistently grouped the populations of humanity into superclusters and clusters, or main branches and sub-branches, that are consistent with the traditional classification of racial divisions and subdivisions. The Caucasian branch on the tree (B) has a main European sub-branch (76) which includes the Lapps or Sami of the Arctic region, then a separate sub-Arctic European sub-branch (75), then a separate Northern European sub-branch (56) which has a northwest European branch (92) represented by the English and German samples.
As seen in the above phylogenetic tree the major genetic divisions or branches of human populations are Africans (A), Caucasians (B), Greater Asians (C), Amerindians (D) and Australopapuans (E). Genetic studies have consistently grouped the populations of humanity into superclusters and clusters, or main branches and sub-branches, that are consistent with the traditional classification of racial divisions and subdivisions. The Caucasian branch on the tree (B) has a main European sub-branch (76) which includes the Lapps or Sami of the Arctic region, then a separate sub-Arctic European sub-branch (75), then a separate Northern European sub-branch (56) which has a northwest European branch (92) represented by the English and German samples.




I’ve been talking with somebody about the concept of race these last couple of days, and I’ve been wondering: Can we properly define what constitutes a different ‘race’ What are the markers and borders that allow us to judge whether a group of humans belong within that category or not? Obviously races can be properly categorized using morphology (different phenotypical features) but can ‘race’ be spotted on the genetic level as well?
“Obviously races can be properly categorized using morphology (different phenotypical features) but can ‘race’ be spotted on the genetic level as well?”
________________________
This is a good question, opponents to racial categorisation want us to point to the proverbial
“negroid” or “asian” gene to “settle” the question.
What they fail to recognise is that coupling negroid adults produce offspring that are like themselves. European coupling adults also produce offspring of their phenotype etc….
Conversely a ‘mixed’ race coupling pair produce ‘mixed’ race offspring.
This is unmistakable and confirms heredity. Naturally we would want to know how that is encapsulated genetically. I’m not aware that a definitive discovery has been made but to my laymans mind it seems an obvious line of enquiry.
But to deny the existence of race is crazy!
The previous response provides perhaps the best answer to give to a layman.
As to the genetic issue, the simple answer is yes, given enough criteria or genetic markers to work with the overlap between races shrinks to zero, enabling forensic geneticists to determine race from a DNA sample as accurately as a forensic physical anthropologist can determine it from skeletal remains, which is to say at least as accurately, if not more, as it can be determined visually from phenotype.
For greater detail there are many sources, especially as your question is closely related to “Lewontin’s fallacy,” which has been the subject of much discussion since 1972 when genetics was not yet advanced enough to provide a definitive answer. By 2008, per Charles Murray, genetics had advanced to the point it could determine a person’s race with 99.9% accuracy:
http://www.unz.com/pfrost/lewontins-fallacy/
Wikipedia provides a necessarily somewhat technical synopsis of “Human Genetic Diversity: Lewontin’s Fallacy,” a 2003 paper by A. W. F. Edwards:
“Edwards argued that while Lewontin’s statements on variability are correct when examining the frequency of different alleles (variants of a particular gene) at an individual locus (the location of a particular gene) between individuals, it is nonetheless possible to classify individuals into different racial groups with an accuracy that approaches 100 percent when one takes into account the frequency of the alleles at several loci at the same time. This happens because differences in the frequency of alleles at different loci are correlated across populations—the alleles that are more frequent in a population at two or more loci are correlated when we consider the two populations simultaneously. Or in other words, the frequency of the alleles tends to cluster differently for different populations.
In Edwards’s words, “most of the information that distinguishes populations is hidden in the correlation structure of the data.” These relationships can be extracted using commonly used ordination and cluster analysis techniques. Edwards argued that, even if the probability of misclassifying an individual based on the frequency of alleles at a single locus is as high as 30 percent (as Lewontin reported in 1972), the misclassification probability becomes close to zero if enough loci are studied.
Edwards’s paper stated that the underlying logic was discussed in the early years of the 20th century. Edwards wrote that he and Luigi Luca Cavalli-Sforza had presented a contrasting analysis to Lewontin’s, using very similar data, already at the 1963 International Congress of Genetics. Lewontin participated in the conference but did not refer to this in his later paper. Edwards argued that Lewontin used his analysis to attack human classification in science for social reasons.”
https://en.wikipedia.org/wiki/Human_Genetic_Diversity:_Lewontin's_Fallacy
The Entire Edwards study is available at: https://onlinelibrary.wiley.com/doi/abs/10.1002/bies.10315
Oy vey, the goyim know about DNA! Shut it down!